Systematiek op z’n informatiekundigs
Pieter Wisse
De biologie telt van oudsher een discipline die systematiek heet. Dat ontdekte ik overigens zojuist pas via het boek Biological Systematics (Addison-Wesley, 1974) door H.H. Ross. Meteen kwam de vraag op, of informatiekundig ontwerpers voor modellering wellicht iets kunnen opsteken van biosystematiek. Of is het soms omgekeerd? In 2012 ben ik met een antwoord wellicht nogal aan de late kant, maar ik herken nergens dat iemand anders mij ermee vóór geweest is. Indien dergelijke verkenningen tot en met uitwerkingen wel degelijk reeds gemaakt zijn, hoop ik dat de beter ingelichte lezer mij daarover inlicht (zodat ik deze tekst in dat opzicht prompt kan verbeteren).
Kenschets van biosystematiek
Volgens Ross zijn er ruwweg twee varianten van biosystematiek. De ene dateert van vóór de evolutietheorie, de andere van erná.
Zolang levensvormen (soorten) ònveranderlijk gedacht waren, bleef systematiek beperkt tot hun, zeg maar, nevengeschikte indeling (plus een hiërarchische classificatie voor overzicht en toegankelijkheid). Synchronisch, zou ik met een term uit de linguïstiek kortweg zeggen. Volgens de aanname van soortelijke continuïteit was oriëntatie op bestaande organismen nodig en voldoende (p. 6): “[S]pecies were defined on the basis of the possession of distinctive characters.”
Hier is het al oppassen. Met karakter(trek) is een eigenschapsoort bedoeld. Zo’n eigenschapsoort kan bij een ¾ exemplaar van een ¾ organisme een bepaalde waarde aannemen. Voor de radicaalste indeling van organismesoorten geldt dan èlk verschil tussen eigenschapwaarden als relevant c.q. grondslag voor aparte soorten.
Een principiële vraag is natuurlijk wat als eigenschapsoorten meetelt en welke bijbehorende eigenschapwaarden in aanmerking komen. Want dat verandert. Bijvoorbeeld met een microscoop raakte zichtbaar wat voordien verborgen gebleven was. Die gewijzigde praktijk leidde tot aangepaste theorie, terwijl omgekeerd een nieuwe theorie aanleiding kan zijn voor ontwikkeling van instrumentarium voor waarneming.
Door de evolutietheorie ging ook en vooral de biosystematiek zich tevens naar de tijddimensie richten. Diachronisch, dus. Welke positie heeft een organismesoort (species) in zoiets als een stamboom? Ofwel, uit welke soort (en wanneer) is een ‘nieuwe’ soort ontstaan en welke verdere soorten zijn daaruit ontstaan (en wanneer), enzovoort. Voor deze fylogenie is het uiteraard extra oppassen met de overgangen tussen de veronderstelde soorten.
Met het accent op afstammingslijnen zijn genetische eigenschapsoorten op de voorgrond getreden ter bepaling van wat al dan niet erkenning krijgt als een organismesoort. Daarvoor oriënteert de (bio)systematiek zich tegenwoordig (dus) sterk op de moleculaire biologie.
Overeenkomst met traditionele objectgerichtheid
Laat ik voorop stellen dat mijn kenschets hierboven stellig een zeer gebrekkige karikatuur van biosystematiek is. Desondanks waag ik me aan enkele verdere opmerkingen. Zo valt mij op, dat het axiomatische element van (zulke) systematiek nog steeds het organisme als exemplaar is. En zo’n exemplaar (lees ook: individu) wordt naar precies één soort geordend (lees ook: tot element gesystematiseerd).
Nu stelt Ross onder de noemer van “ontogenetic changes” dat (p. 25) “[i]n all individuals, characters and patterns of characters change during development.” Dit roept uiteraard de vraag op, wat (nog) telt als hetzelfde individu. Over eigenschapsoorten doet Ross in elk geval niet moeilijk (p. 26): “The characters of the individual embrace the sum total of all characters of all stages.” Voor die allesomvattende verzameling bestaat blijkbaar ook een naam, “holomorphology.”
De praktische modelleeropgave is wèl moeilijk, althans volgens de ¾ vergaand impliciete ¾ veronderstellingen die Ross laat gelden voor biosystematiek. Nota bene, die zijn vergelijkbaar met zgn objectgerichtheid voor informatievoorziening. Als louter opslagplaats van “the sum total of all characters of all stages” is een (informatie)object nog voorstelbaar. Maar het doel met zo’n object is om informatie selectief beschikbaar te stellen. Dat vergt structuur, die echter met zomaar “the sum total” juist ... totaal ontbreekt.
Die structuur is met nadruk nodig ter ordening van “all characters.” Wat beweert Ross daar? Gaat hij uit van een beperkte, gecontroleerde verzameling met eigenschapsoorten? Of hoogstens van verschillende van die verzamelingen, zgn codes, voor bacteriën, dieren en planten? Daar heeft het veel van weg. En, ja, de eigenschapsoorten volgens die gesloten, formeel gestandaardiseerde verzamelingen passen wellicht (nog) allemaal in één of ander informatiekundig objectsoort.
Systematiek ... verstelselmatigd met Metapatroon
Zodra de grens van een ènkele discipline wegvalt, is de verzameling van “all characters” echter veranderlijk, praktisch oneindig. Dan is expliciete structuur onontbeerlijk. En óók die structuur moet aanpasbaar zijn! De traditionele objectgerichtheid voorziet daarin niet. Dat lukt wel met verbijzondering volgens relevante contexten. De methode die ik daarvoor ontwierp heet Metapatroon (Engels: Metapattern).
Voor een eerste indruk presenteer ik hier op z’n Metapatroons een informatiemodel voor gestructureerde holomorfologie. Ik wijs erop dat figuur 1 nog lang geen stelselmatig model biedt. Zo ontbreken de instellingen die de respectievelijke classificaties certificeren e.d. (en als zodanig de context ervoor vormen).
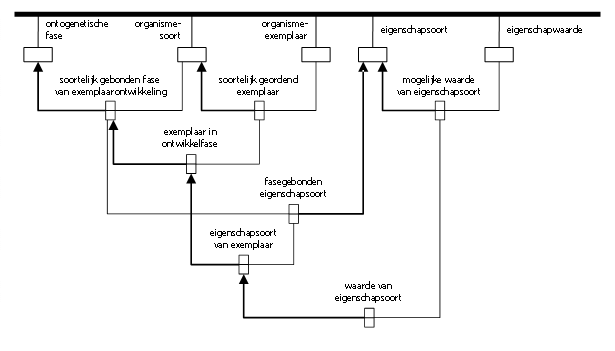
figuur 1: een begin met gestructureerde holomorfologie.
Merk op dat het Metapatroonmodel de positionering van eigenschappen als het ware binnenstebuiten keert. Informatie over de relevante waarden is niet ongestructureerd ‘in’ het organisme-exemplaar in kwestie gedacht, maar opzettelijk èn expliciet gestructureerd via getrapte contextualiseringen.
Perspectief voor systematiek
Aldus nodigt Metapatroon m.i. zelfs uit om de overgeleverde aannames van/voor de biosystematiek te heroverwegen. Omdat Metapatroon samenhang tussen verschillen laat vestigen, kunnen voorheen apart veronderstelde voorwaarden losser gesteld worden; dankzij verband blijft structuur immers overeind, zelfs steviger. Bijvoorbeeld, juist door eigenschappen niet opeen te hopen in het individu, kan dat individu zowel syn- als diachronisch meervoudig worden geordend/gesystematiseerd. Het hangt er maar vanaf. Pèr context heeft het individu desbetreffende eigenschappen (soorten met waarden), terwijl samenhang bestaat tussen aldus verbijzonderde gedragsvormen van het tegelijk als continu veronderstelde individu. Voor contextualiseringen komen dan uiteraard in aanmerking wat Ross nog vergaand ongestructureerd beschrijft als differentiërende factoren, te weten geografie en (overige) ecologie. Met ecologie als integraal en integrerend onderdeel van ordening van organismesoorten verkrijgt systematiek het vitale interdependente karakter.
Maart 2012, webeditie 2012 © Pieter Wisse (Information Dynamics)
Publicaties over Metapatroon zijn toegankelijk via Metapatroon, handboek stelselmatig informatieverkeer.